Hãy nhập câu hỏi của bạn vào đây, nếu là tài khoản VIP, bạn sẽ được ưu tiên trả lời.

Bài 1:
- Một chuỗi thức ăn gồm nhiều loài có quan hệ dinh dưỡng với nhau và mỗi loài là một mắt xích của chuỗi. Trong một chuỗi, một mắt xích vừa ăn thịt mắt xích phía sau, vừa bị mắt xích phía trước ăn thịt.
Ví dụ: cỏ —> thỏ—»cáo
- Lưới thức ăn được hình thành từ nhiều chuỗi thức ăn trong quần xã. Trong một lưới thức ăn một loài sinh vật không phải chỉ tham gia vào một chuỗi thức ăn mà còn tham gia đồng thời vào các chuỗi thức ăn khác, hình thành nên nhiều mắt xích chung tất cả các chuỗi thức ăn với nhiều mắt xích chung hợp thành một lưới thức ăn.
Có hai loại chuỗi thức ăn:
Chuỗi thức ăn được mở đầu bằng cây xanh, sau đến động vật ăn thực vật và tiếp nữa là các loài động vật ăn động vật.
Ví dụ: Cây ngô -» sâu ăn lá ngô —> nhái —> rắn hổ mang —» diều hâu.
Chuỗi thức ăn mở đầu bằng chất hữu cơ bị phân giải, sau đến các loài động vật ăn thịt.
Ví dụ: Lá, cành cây khô —» mối -» nhện —> thằn lằn.
Bài 2 :
- Ví dụ về các bậc dinh dưỡng cùa 1 quần xã tự nhiên (quần xã đồng cỏ):
+ Sinh vật sản xuất: cây cỏ, cây bụi.
+ Sinh vật tiêu thụ bậc 1: sâu ăn lá cây, rệp, chuột.
+ Sinh vật tiêu thụ bậc 2: chim sâu, rắn.
+ Sinh vật tiêu thụ bậc cao nhất: diều hâu.
+ Sinh vật phân huỷ: vi khuẩn, nấm, mối, giun đất.
- Vi dụ về các bậc dinh dưỡng cùa 1 quần xã tự nhiên (quần xã suối):
+ Sinh vật sản xuất: tảo lục, tảo silic, thuỷ tức.
+ Sinh vật tiêu thụ bậc 1: muỗi nước, tôm, cá mè.
+ Sinh vật tiêu thụ bậc 2: nhện nước, cá quả.
+ Sinh vật phân huỷ: vi khuẩn, giun.
+ Chất hữu cơ từ ngoài theo dòng suối: các mẩu lá cây, cành cây, rác,...
- Ví dụ về bậc dinh dưỡng của quần xã nhân tạo (quần xã đồng lúa):
+ Sinh vật sản xuất: cây lúa.
+ Sinh vật tiêu thụ bậc 1: sâu đục thân lúa, rệp. chuột.
+ Sinh vật tiêu thụ bậc 2: chim sâu, rắn.
+ Sinh vật tiêu thụ bậc cao nhất: diều hâu.
+ Sinh vật phân huỷ: vi khuẩn, nấm, giun đất.
Trên mỗi phân tử mARN thường có một số riboxom cùng hoạt đọng được gọi là poliriboxom. sự hình thành poliriboxom là sau khi riboxom thứ nhất dịch chuyển được 1 đoạn thì riboxom thứ 2 liên kết vào mARN.
Tiếp theo đó là riboxom thứ 3, 4 .... Như vậy, mỗi 1 phân tử mARN có thể tổng hợp từ 1 đến nhiều chuỗi polipeptit cùng loại rồi tự hủy. Các riboxom được sử dụng qua vài thế hệ tế bào và có thể tham gia vào tổng hợp bất cứ loại protein nào.
Các thành phần tham gia vào quá trình dịch mã phân tử protein gồm có: mARN, t ARN, riboxoom, axit amin và ATP.
ADN là mạch khuôn tổng hợp mARN, thông qua mARN truyền thông tin quy định tổng hợp protein.
ADN gián tiếp tham gia vào quá trình tổng hợp phân tử protein.
Chọn D
Câu 5: Hãy chọn phương án đúng
Quy luật phân li độc lập thực chất nói về
A. Sự phân li độc lập của các tính trạng
B. Sự phân li kiểu hình theo tỉ lệ 9 : 3 : 3 : 1
C. Sự tổ hợp của các alen trong quá trình thụ tinh
D. Sự phân li độc lập của các alen trong quá trình giảm phân.
* Diễn biến – Khi enzim ARN pôlimeraza bám vào promoter (vùng khởi đầu của gen) làm gen tháo xoắn và tách 2 mạch đơn, làm lộ ra mạch khuôn (mạch gốc) (mạch có chiều 3‘ 5‘) – Enzim ARN pôlimeraza di chuyển dọc theo mạch gốc và gắn các nuclêôtit tự do trong môi trường nội bào liên kết với các nuclêôtit trên mạch khuôn theo nguyên tắc bổ sung (U-A, A-T; G-X, X-G) tạo nên phân tử mARN theo chiều 5‘ 3‘ – Khi enzim di chuyển đến cuối gen gặp tín hiệu kết thúc thì dừng quá trình phiên mã và phân tử mARN vừa được tổng hợp được giải phóng. Vùng nào trên gen phiên mã xong thì đóng xoắn. * Kết quả: ở tế bào nhân sơ mARN sau khi tổng hợp ở dạng trưởng thành và trực tiếp làm khuôn tổng hợp chuỗi pôlipeptit. Còn ở tế bào nhân thực mARN sau khi tổng hợp ở dạng sơ khai, sau đó được cắt bỏ các đoạn intron, nối các đoạn exon lại với nhau thành mARN trưởng thành, đi qua màng nhân ra tế bào chất làm khuôn tổng hợp chuỗi pôlipeptit.
* Diễn biến – Khi enzim ARN pôlimeraza bám vào promoter (vùng khởi đầu của gen) làm gen tháo xoắn và tách 2 mạch đơn, làm lộ ra mạch khuôn (mạch gốc) (mạch có chiều 3‘ 5‘) – Enzim ARN pôlimeraza di chuyển dọc theo mạch gốc và gắn các nuclêôtit tự do trong môi trường nội bào liên kết với các nuclêôtit trên mạch khuôn theo nguyên tắc bổ sung (U-A, A-T; G-X, X-G) tạo nên phân tử mARN theo chiều 5‘ 3‘ – Khi enzim di chuyển đến cuối gen gặp tín hiệu kết thúc thì dừng quá trình phiên mã và phân tử mARN vừa được tổng hợp được giải phóng. Vùng nào trên gen phiên mã xong thì đóng xoắn. * Kết quả: ở tế bào nhân sơ mARN sau khi tổng hợp ở dạng trưởng thành và trực tiếp làm khuôn tổng hợp chuỗi pôlipeptit. Còn ở tế bào nhân thực mARN sau khi tổng hợp ở dạng sơ khai, sau đó được cắt bỏ các đoạn intron, nối các đoạn exon lại với nhau thành mARN trưởng thành, đi qua màng nhân ra tế bào chất làm khuôn tổng hợp chuỗi pôlipeptit.
- Sự hoạt động cùa opêron chịu sự điều khiển của một gen điều hoà (regulator: R) nằm ở trước opêron.
- Bình thường, gen R tổng hợp ra một loại prôtêin ức chế gắn vào gen chỉ huy do đó gen cấu trúc ở trạng thái bị ức chế nên không hoạt động. Khi có chất cảm ứng (ví dụ lactôzơ) thì opêron chuyển sang trạng thái hoạt động (cảm ứng).
- Biểu hiện ở gen R và opêron Lac trong trạng thái ức chế:
+ Trong điều kiện bình thường (môi trường không có chất cảm ứng: đường lactôzơ), gen điều hoà (R) phiên mã tạo ra mARN của nó, mARN này được sử dụng để tổng hợp ra chất ức chế (prôtêin ức chế).
+ Chất ức chế đến bám vào gen chỉ huy. Gen chỉ huy bị ức chế do đó các gen cấu trúc không phiên mã.
- Biểu hiện ở gen R và opêron Lac khi có chất cảm ứng lactôzơ:
+ Khi môi trường nuôi E. coli có đường lactôzơ (đặc biệt là môi trường trước đó không có lactôzơ và thiếu glucôzơ) thì lactôzơ tác dụng với chất ức chế, chất ức chế bị bất hoạt. Do vậy, nó không còn có thể kết hợp với gen chỉ huy nữa.
+ Gen chỉ huy được tự do điều khiển quá trình phiên mã của cả opêron, mARN cùa các gen A. B, C được tổng hợp và sau đó được sử dụng để dịch mã tổng hợp các prôtêin enzim tương ứng. Đó là trạng thái cảm ứng (hoạt động) của opêron.
- Khi lactôzơ bị phân giải hết, chất ức chế được giải phóng, chất ức chế chuyển từ trạng thái bất hoạt sang trạng thái hoạt động đến bám vào gen chỉ huy và opêron lại chuyển sang trạng thái bị ức chế.
- Sự hoạt động cùa opêron chịu sự điều khiển của một gen điều hoà (regulator: R) nằm ở trước opêron.
- Bình thường, gen R tổng hợp ra một loại prôtêin ức chế gắn vào gen chỉ huy do đó gen cấu trúc ở trạng thái bị ức chế nên không hoạt động. Khi có chất cảm ứng (ví dụ lactôzơ) thì opêron chuyển sang trạng thái hoạt động (cảm ứng).
- Biểu hiện ở gen R và opêron Lac trong trạng thái ức chế:
+ Trong điều kiện bình thường (môi trường không có chất cảm ứng: đường lactôzơ), gen điều hoà (R) phiên mã tạo ra mARN của nó, mARN này được sử dụng để tổng hợp ra chất ức chế (prôtêin ức chế).
+ Chất ức chế đến bám vào gen chỉ huy. Gen chỉ huy bị ức chế do đó các gen cấu trúc không phiên mã.
- Biểu hiện ở gen R và opêron Lac khi có chất cảm ứng lactôzơ:
+ Khi môi trường nuôi E. coli có đường lactôzơ (đặc biệt là môi trường trước đó không có lactôzơ và thiếu glucôzơ) thì lactôzơ tác dụng với chất ức chế, chất ức chế bị bất hoạt. Do vậy, nó không còn có thể kết hợp với gen chỉ huy nữa.
+ Gen chỉ huy được tự do điều khiển quá trình phiên mã của cả opêron, mARN cùa các gen A. B, C được tổng hợp và sau đó được sử dụng để dịch mã tổng hợp các prôtêin enzim tương ứng. Đó là trạng thái cảm ứng (hoạt động) của opêron.
- Khi lactôzơ bị phân giải hết, chất ức chế được giải phóng, chất ức chế chuyển từ trạng thái bất hoạt sang trạng thái hoạt động đến bám vào gen chỉ huy và opêron lại chuyển sang trạng thái bị ức chế.
* Cách li trước hợp tử
- Cách li nơi ở (sinh cảnh): Mặc dù sống trong cùng một khu vực địa lí nhưng những cá thể của các loài có họ hàng gần gũi nhưng sống ở những sinh cảnh khác nhau nên không thể giao phối với nhau.
- Cách li tập tính: Các cá thể của các loài khác nhau có thể có những tập tính giao phối riêng nên giữa chúng thường không giao phối với nhau.
- Cách li thời gian (mùa vụ): Các cá thể thuộc các loài khác nhau có thể sinh sản vào những mùa khác nhau nên chúng không có điều kiện giao phối với nhau.
- Cách li cơ học: Các cá thể thuộc các loài khác nhau có thể có cấu tạo các cơ quan sinh sản khác nhau nên chúng không thể giao phối với nhau. Ví dụ, các cây khác loài có cấu tạo hoa khác nhau nên hạt phấn của loài cây này không thể thụ phấn cho hoa của loài cây khác.
* Cách li sau hợp tử
Cách li sau hợp tử là những trở ngại ngăn cản việc tạo ra con lai hoặc ngăn cản việc tạo ra con lai hữu thụ.
Trả lời: . -
* Cách li trước hợp tử
- Cách li nơi ở (sinh cảnh): Mặc dù sống trong cùng một khu vực địa lí nhưng những cá thể của các loài có họ hàng gần gũi nhưng sống ở những sinh cảnh khác nhau nên không thể giao phối với nhau.
- Cách li tập tính: Các cá thể của các loài khác nhau có thể có những tập tính giao phối riêng nên giữa chúng thường không giao phối với nhau.
- Cách li thời gian (mùa vụ): Các cá thể thuộc các loài khác nhau có thể sinh sản vào những mùa khác nhau nên chúng không có điều kiện giao phối với nhau.
- Cách li cơ học: Các cá thể thuộc các loài khác nhau có thể có cấu tạo các cơ quan sinh sản khác nhau nên chúng không thể giao phối với nhau. Ví dụ, các cây khác loài có cấu tạo hoa khác nhau nên hạt phấn của loài cây này không thể thụ phấn cho hoa của loài cây khác.
* Cách li sau hợp tử
Cách li sau hợp tử là những trở ngại ngăn cản việc tạo ra con lai hoặc ngăn cản việc tạo ra con lai hữu thụ.
Quá trình dịch mã tại ribôxôm chia thành 2 giai đoạn: – Hoạt hóa axit amin Trong tế bào chất, nhờ enzim đặc hiệu và ATP, mỗi axit amin được hoạt hóa và gắn với tARN tương ứng tạo nên phức hợp axit amin – tARN (aa – tARN). – Tổng hợp chuỗi pôlipeptit. Gồm 3 giai đoạn + Mở đầu Tiểu đơn vị bé của ribôxôm gắn với mARN ở vị trí đặc hiệu nằm gần côđon mở đầu. Bộ ba đối mã của phức hợp mở đầu Met – tARN (UAX) khớp bổ sung với côđon mở đầu (AUG) trên mARN. Tiểu đơn vị lớn của ribôxôm kết hợp tạo ribôxôm hoàn chỉnh sẵn sàng tổng hợp chuỗi pôlipeptit. + Kéo dài chuỗi pôlipeptit Anticôđon của phức hợp thứ 2 vào gắn bổ sung với côđon thứ 2 trên mARN, aa thứ 2 liên kết với aa Met (trước) bằng liên kết peptit. Ribôxôm lại dịch chuyển đi một côđon trên mARN và cứ tiếp tục như vậy đến cuối mARN. + Kết thúc Khi ribôxôm tiếp xúc với mã kết thúc trên mARN thì quá trình dịch mã hoàn tất. Nhờ enzim đặc hiệu, axit amin mở đầu được cắt khỏi chuỗi pôlipeptit vừa tổng hợp. Chuỗi pôlipeptit tiếp tục hình thành các cấu trúc bậc cao hơn, trở thành prôtêin có hoạt tính sinh học. Trong quá trình dịch mã , mARN không gắn với từng ribôxôm riêng rẽ mà đồng thời gắn với một nhóm ribôxôm gọi là pôliribôxôm (gọi tắt là pôlixôm) giúp tăng hiệu suất tổng hợp prôtêin.
Câu 6: Hãy chọn phương án đúng.
Vai trò của enzim ADN polymelaza trong quá trình nhân đôi ADN là:
A. Tháo xoắn phân tử ADN
B. Bẻ gãy các liên kết hidro giữa hai mạch ADN
C. Lắp ráp các nucleotit tự do theo nguyên tắc bổ sung với mỗi mạch khuôn của ADN
D. Cá A, B, C
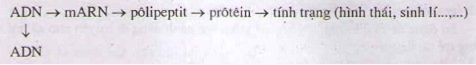

Chọn B